
(Paper presented at the World Textile Congress on Natural and Natural-Polymer Fibres, 1997, University of Huddersfield, UK)
Animal hair fibres are based on cross-linked proteins known as keratins. The major fibre in this group is sheep’s wool but other fibres of considerable commercial importance are those obtained from animals such as goats (cashmere and mohair), the bactrian camel and the South American camelids. The latter are known as speciality or luxury fibres due to their softness, lustre and scarcity relative to other fibres.
Identification of both raw and processed speciality animal fibres is important to help combat adulteration or false declaration and ensure adherence to international trading agreements. Cashmere, in particular, has frequently been found to be adulterated with much cheaper fibres such as sheep’s wool or yak hair. New legal pressures concerning garment labelling requirements within the EC underline the need for exact analytical methods but these can be difficult to achieve, particularly for materials of biological origin.
To date, the most important means of identifying animal fibres have been light and scanning electron microscopy. Chemically there is little difference between cashmere, wool and yak hair. Therefore, microscopists have to rely on physical parameters such as internal morphology, fibre diameter and cuticle scale height. Detailed measurements have to be made on hundreds of individual fibres for each sample received for analysis. In addition, there are some fibre combinations that even an experienced microscopist cannot differentiate (e.g. fine yak hair and brown cashmere; the dark pigmentation of these fibres prevents assessment of their internal structure). Clearly, there is the need for a quick and objective method of fibre analysis.
This article summarises a programme of work carried out at BTTG to develop novel analytical techniques for fibre identification based on recent advances in molecular biology.
Location of DNA in fibres
Animal hair fibres are constructed from cellular material. In the living cells of plants and animals DNA (deoxyribonucleic acid), the chemical molecule that carries the hereditary information, is contained inside spherical bodies known as nuclei. Nuclei contained within immature hair follicles can be directly visualised using the technique of in situ DNA hybridisation and a high-powered microscope. As the follicle matures, the nuclei become elongated but the signal does not extend into the more mature parts of the fibre where keratinisation has taken place (Figure 1). During the process of keratinisation the cells become metabolically inactive, however remnants of the nuclear material containing DNA remain trapped inside the fibre. The recalcitrant and waterproof nature of fibres helps to protect the DNA from chemical and biological degradation.
In recent years ‘ancient DNA’ has been recovered and analysed from the remains of plants and animals that are thousands of years old. Work at DWI (Aachen, West Germany) and BTTG (Manchester, UK) has confirmed that DNA can also be recovered from animal hair shafts (Kalbe et al, 1988; Hamlyn et al, 1990). This has led to the development of DNA fibre profiling techniques at BTTG for the specific identification of wool and yak hair.
Extraction of DNA and analysis of raw fibres
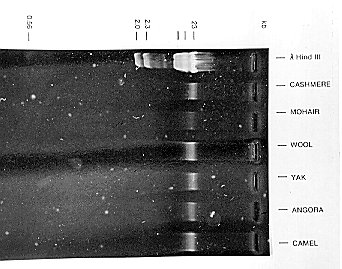
Various methods have been developed for the recovery of DNA from keratin fibres. DNA has been extracted from cuticle cells removed from the surface of fibres by washing in an aqueous solution of sodium dodecyl sulphate (Table 1, Figure 2), from cryogenically milled fibres and from whole fibre snippets. For raw fibre analysis, cuticle extraction has the advantage that a large quantity of fibre (typically 20 grams) can be processed yielding sufficient DNA to allow for the detection of the major components in a blend using hybridisation analysis.
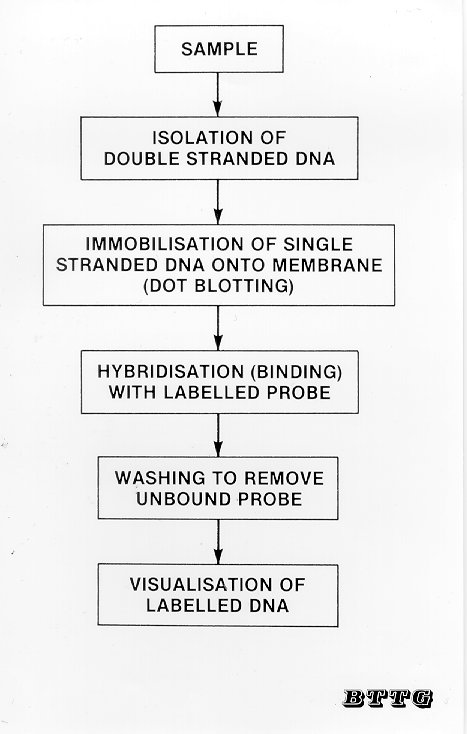
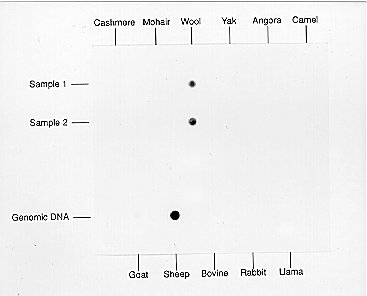
DNA hybridisation analysis of fibres is based on the use of short DNA sequences known as oligonucleotides that are unique to each species. Species-specific oligonucleotides are available that can be used to distinguish between DNA samples from closely related species such as sheep and goats (Hamlyn et al, 1992; Nelson et al, 1992). The analysis is carried out using a simple dot-blot technique (Figure 3). DNA isolated from fibre samples together with control samples of genomic DNA is denatured by boiling to convert the native double-stranded DNA molecules into single strands and then spotted onto a nylon membrane (Bio-Rad). A single-stranded preparation of the oligonucleotide to be used as the probe is then incubated with the membrane to allow hybridisation to take place between the probe and any complementary strands of DNA present in the sample. After free unhybridised probe has been removed by washing the membrane several times, the hybrid double-stranded DNA molecules can be visualised by several different methods according to how the probe has been labelled (i.e. by using radioactive, colorimetric or luminescent labels). For example, a sheep-specific oligonucleotide probe can be used to detect the presence of wool but does not give a positive signal with DNA extracted from other types of fibre (Figure 4).
Analysis of processed materials
Although standard hybridisation methods can be used for the identification of samples of raw fibres, with scoured fibres and dyed garments because much of the DNA has been degraded and removed during processing, it is not possible to use conventional methods of DNA analysis. However, the development of in vitro DNA amplification technology known as the polymerase chain reaction (PCR) allows for the analysis of minute quantities of DNA. In essence, PCR reproduces what happens when the DNA inside a cell is replicated during cell division. By using an automated temperature cycling block (Figure 5) this ‘enzymatic copying system’ can be used to produce millions of identical copies of a specified DNA sequence after only 1-2 hours (Figure 6). These copies provide sufficient DNA for further analysis (e.g. sequencing), or can simply confirm the presence of the target sequence in the sample (Figure 7). Thus, if species-specific primers are employed the presence or absence of an adulterant can be quickly determined and this result can later be confirmed by DNA sequencing. The resulting DNA sequence can be compared to published sequences providing courts with evidence of an exact match that leaves no room for doubt. Because of the prodigious sensitivity of PCR it is essential that negative controls are included with the samples and go through the same extraction process.
Using this approach, DNA has been successfully amplified from samples of dyed material that did not yield detectable levels of DNA using conventional extraction techniques and we have shown that PCR is capable of amplifying and therefore allowing rapid detection of minor components in mixed DNA samples (Hamlyn et al, 1996). Although this type of analysis is not quantitative it is a simple and rapid test providing useful criteria for the confirmation of fraudulent substitution.
Possibilities for quantitative fibre analysis
To carry out quantitative analysis it is necessary to identify individual fibre shafts. The development of such technology will enable accurate determination of the composition of unknown fibre blends. Recent work has indicated that in situ DNA analysis of single fibre shafts is not practically possible since the DNA is encapsulated inside the waterproof environment of the keratinised cells (Broadbent, 1997). Attempts to get at this DNA leads to the dissolution of the fibres. Therefore extraction-based techniques will have to be employed. A key strategy will be to improve the efficiency of recovery of DNA from whole fibres. Proteolytic enzymes such as proteinase K (Sigma) can partially degrade animal fibres but the cells of the cortex remain intact even after several days incubation with the enzyme (Figure 8). Other enzyme systems are currently being evaluated. Preliminary investigations have indicated that DNA can be extracted and amplified from single fibres of raw yak hair treated with the enzyme papain (Hamlyn, unpublished).
A second problem relates to the large number of fibres that will have to be analysed to give an accurate result making any test prohibitively costly using manual techniques. However, automated systems are being developed for both DNA extraction and amplification and the advent of chip-based systems will ultimately lead to low cost procedures that can handle large numbers of samples.
REFERENCES
Broadbent, E.L. (1997). The identification and quantification of the constituents in processed animal fibre blends. Ph.D. thesis, University of Leeds.
Hamlyn, P.F., Nelson, G., and McCarthy, B.J. (1990). Applied molecular genetics - new tools for animal fibre identification, in, Speciality Fibres: Scientific, Technological and Economical Aspects (A. Körner, F.J. Wortmann, G. Wortmann and H. Höcker, eds.), Schriftenreihe des Deutschen Wollforschungsinstitutes, a.d. Technischen Hochschule Aachen, e.v.,106, pp. 249-258.
Hamlyn, P.F., Nelson, G., and McCarthy, B.J. (1992) Wool Fibre Identification using Novel Species-Specific DNA Probes. Journal of the Textile Institute 83(1), 97-103.
Hamlyn, P.F., Ramsbottom, S., McCarthy, B.J. & Nelson, G. (1996). Analysis of speciality fibres using DNA amplification techniques, in, Metrology and Identification of Speciality Animal Fibres (J.P. Laker and F.J. Wortman, eds.), European Fine Fibre Network, Occasional Publication No. 4, Macaulay Land Use Research Institute, Aberdeen, Scotland, pp. 59-68.
Kalbe, J., Kuropka, R., Meyer-Stork, S., Sauter, S.L., Loss, P., Henco, K., Riesner, D., Höcker, H. and Berndt, H. (1988). Isolation and characterisation of high - molecular mass DNA from hair shafts. Biol. Chem. Hoppe-Seyler, 369, 413-416.
Nelson, G., Hamlyn, P.F., and Holden, L. (1992) A Species-Specific DNA Probe for Goat Fibre Identification. Textile Research Journal, 62(10), 590-595.
Table 1 Recovery of DNA from samples of cashmere, wool and yak hair using the cuticle extraction method (note that much lower quantities of DNA are obtained from commercially scoured samples).
|
Fibre Sample |
Treatment |
DNA Yield (ng per gram of sample) |
|
White Cashmere |
Raw * |
75.0 |
|
White Cashmere |
Scoured |
2.5 |
|
White Cashmere |
Scoured |
1.2 |
|
Brown Cashmere |
Scoured |
5.0 |
|
Brown Cashmere |
Scoured |
1.2 |
|
Wool |
Raw * |
50.0 |
|
Yak Hair |
Raw * |
5.0 |
|
Yak Hair |
Scoured |
<1.2 |
*
Raw fibre samples are given a mild laboratory scour prior to cuticle isolation.Figure 1 Visualisation of the nuclei inside a hair follicle using the technique of in situ hybridisation. From Broadbent (1997).
Figure 2 DNA extracted from the cuticle of speciality fibres and visualised by UV fluorescence after resolution on an agarose gel. From Nelson (1992).
Figure 3 Analysis of DNA by the dot-blotting technique.
Figure 4 Hybridisation of a sheep-specific oligonucleotide probe to DNA extracted from various animal fibres. A positive signal was only obtained with wool DNA. From Hamlyn (1992).
Figure 5 Thermal cycling machine used for carrying out the polymerase chain reaction.
Figure 6 Amplification of DNA using the polymerase chain reaction. The primers are synthetically prepared oligonucleotides having sequences complementary to the DNA on either side of the segment of DNA to be amplified.
Figure 7 DNA amplification products from wool, yak hair and cashmere obtained using different primer sets. From Hamlyn (1995).
Figure 8 Degradation of wool after extended incubation with proteinase K. The spiked shaped fragments are released cells of the cortex. From Broadbent (1997).
Copyright © 1997 Paul F Hamlyn
All rights reserved
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}